
慳悈惈傾儈僲巁巆婎偑廤拞偡傞C枛抂懁椞堟偑丄帀幙枌偵懳偟偰崅偄恊榓惈傪帵偟傑偡丅
傾儖僣僴僀儅乕昦偼丄擣抦婡擻忈奞偲婰壇椡偺掅壓傪敽偆恑峴惈拞悤恄宱曄惈幘姵偱偡丅尰嵼丄悽奅偱2600枩恖傪挻偊傞姵幰偑偄傞偲悇掕偝傟偰偍傝丄梊杊朄傗崻杮揑側帯椕朄偺奐敪偑嫮偔朷傑傟偰偄傑偡丅 傾儖僣僴僀儅乕昦姵幰偺擼偵尒傜傟傞昦棟妛揑摿挜偱偁傞傾儈儘僀僪斄偼丄栺40傾儈僲巁巆婎偐傜側傞儁僾僠僪偱偁傞傾儈儘僀僪兝乮A兝丄恾侾乯偑暘巕娫夛崌偟偰惗偠偨慄堐忬嬅廤懱乮傾儈儘僀僪慄堐乯傪庡側惉暘偲偟傑偡丅旕夛崌忬懺偺A兝儌僲儅乕偼柍奞偱偡偑丄暘巕娫夛崌偟偰僆儕僑儅乕壔丄傑偨偼慄堐壔偟偨A兝偼攟梴嵶朎偵懳偟偰撆惈傪帵偟傑偡丅
|
| 恾侾丏A兝1-40偺傾儈僲巁攝楍 慳悈惈傾儈僲巁巆婎偑廤拞偡傞C枛抂懁椞堟偑丄帀幙枌偵懳偟偰崅偄恊榓惈傪帵偟傑偡丅 |
峔憿惗暔妛揑側娤揰偐傜偼丄傾儖僣僴僀儅乕昦偼僞儞僷僋幙偺僐儞儂儊乕僔儑儞曄壔傪宊婡偲偟偰堷偒婲偙偝傟傞乽僐儞儂儊乕僔儑儞昦乿偲憤徧偝傟傞昦婥偺傂偲偮偲偟偰懆偊傞偙偲偑偱偒傑偡丅懄偪丄A兝儌僲儅乕偺悈梟塼拞偵偍偗傞峔憿偼晄婯懃偱偡偑丄暘巕娫夛崌偟偰慄堐偲側傞偨傔偵偼丄A兝偼兝僔乕僩峔憿傪宍惉偡傞昁梫偑偁傝傑偡丅偙偺偨傔丄A兝偺僐儞儂儊乕僔儑儞曄壔偺尨場偲儊僇僯僘儉傪棟夝偡傞偙偲偼丄傾儖僣僴僀儅乕昦偺梊杊朄傑偨偼崻杮揑側帯椕朄傪奐敪偡傞忋偱嬌傔偰廳梫偱偡丅
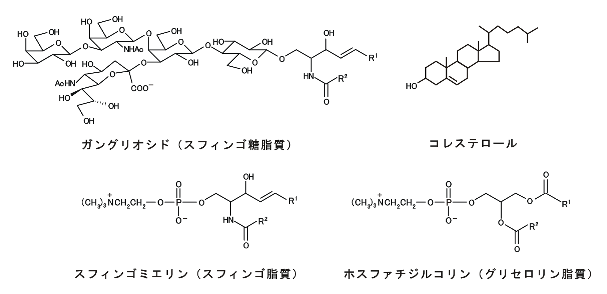
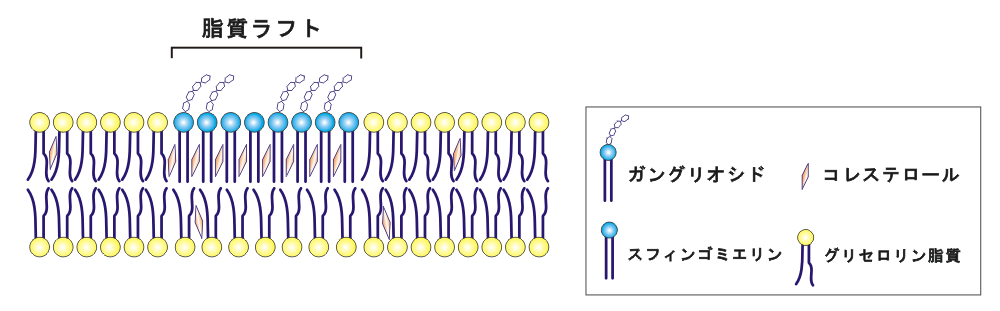
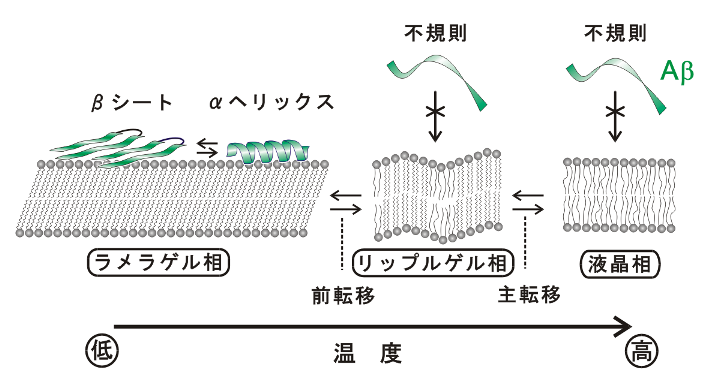
A兝嬅廤偺尨場偲側傞壜擻惈偑巜揈偝傟偰偄傞惗懱惉暘偲偟偰丄嵶朎枌偑偁傝傑偡丅嵶朎枌偺奜梩偵偼丄僈儞僌儕僆僔僪丄僗僼傿儞僑儈僄儕儞丄偍傛傃僐儗僗僥儘乕儖偵晉傓帀幙儔僼僩偲屇偽傟傞枌僪儊僀儞偑丄廃埻偺僌儕僙儘儕儞帀幙偵晉傓枌偐傜暘棧偟偨忬懺偱懚嵼偡傞偲峫偊傜傟偰偄傑偡乮恾俀丄俁乯丅嬤擭丄A兝偑嵶朎枌偺丄摿偵帀幙儔僼僩偵廤愊偟偰暘巕娫夛崌偡傞偙偲偵傛傝丄傾儈儘僀僪慄堐妀乮拻宆乯傪宍惉偡傞壜擻惈偑懡偔偺尋媶偵傛傝帵偝傟偰偄傑偡丅 僋儘僀僣僼僃儖僩丒儎僐僽昦側偳偺尨場偲側傞僾儕僆儞僞儞僷僋幙偺嬅廤傕嵶朎枌偺帀幙儔僼僩乮傑偨偼儔僼僩偲椶帡偡傞帀幙慻惉傪帩偮枌僪儊僀儞偱偁傞僇儀僆儔乯偱惗偠傞偲峫偊傜傟偰偄傞偙偲偐傜丄帀幙儔僼僩偵偼僞儞僷僋幙偺嬅廤傗昦尨宆峔憿傊偺揮堏傪懀偡惈幙偑偁傞偲巚傢傟傑偡丅
 |
| 恾俀丏帀幙暘巕偺峔憿 嵶朎枌傪峔惉偡傞僗僼傿儞僑帀幙偺応崌丄傾僔儖嵔乮R乯偺傎偲傫偳偼朞榓扽壔悈慺嵔偱偁傝丄峝偄枌傪嶌傝傗偡偄丅偙傟偵懳偟偰丄僌儕僙儘儕儞帀幙偺傾僔儖嵔偼丄堦斒偵cis宆偺擇廳寢崌傪娷傒丄廮傜偐偄枌傪嶌傞丅 |
 |
| 恾俁丏帀幙儔僼僩 僗僼傿儞僑儈僄儕儞傗僐儗僗僥儘乕儖偵晉傓帀幙儔僼僩偲屇偽傟傞枌僪儊僀儞偑丄廃埻偺僌儕僙儘儕儞帀幙偵晉傓枌偐傜暘棧偟偨忬懺偱懚嵼偡傞偲峫偊傜傟偰偄傞丅儔僼僩撪偺帀幙暘巕偼丄斾妑揑枾偵僷僢僉儞僌偝傟偨丄拋彉崅偄忬懺偵側偭偰偄傞丅 |
儔僼僩奜偺嵶朎枌偼丄傾僔儖嵔拞偵僔僗宆偺擇廳寢崌傪帩偮僌儕僙儘儕儞帀幙傪懡偔娷傓偨傔丄塼徎憡乮柍拋彉塼懱憡乯偺廮傜偐偄枌偱偁傞偺偵懳偟丄儔僼僩撪偼帀幙暘巕偺僷僢僉儞僌偑枾偱偁傞拋彉塼懱憡偲屇偽傟傞峝偄枌偺忬懺偵側偭偰偄傑偡丅 摉尋媶幒偱偼丄枌偺憡忬懺偺堘偄偑丄A兝偺儔僼僩傊偺廤愊偍傛傃暘巕娫夛崌偺尨場偲側傞壜擻惈偵拝栚偟丄儁僾僠僪偲帀幙枌偺憡屳嶌梡傪庬乆偺暘岝庤朄傪梡偄偰尋媶傪峴側偭偰偄傑偡丅
嵶朎枌偺庡梫側惉暘偱偁傞儂僗僼傽僠僕儖僐儕儞乮PC乯偺帀幙擇廳枌偼崅壏偱塼徎憡偱偁傝丄壏搙掅壓偵傛傝儕僢僾儖僎儖憡傪宱偰儔儊儔僎儖憡偵揮堏偟傑偡乮恾係乯丅塼徎憡偲儕僢僾儖僎儖憡偺娫偺憡揮堏傪庡揮堏丄儕僢僾儖僎儖憡偲儔儊儔僎儖憡偺娫偺憡揮堏傪慜揮堏偲屇傃傑偡丅 A兝偺僠儘僔儞巆婎偺寀岝傪棙梡偟偰丄A兝偲PC帀幙擇廳枌偺憡屳嶌梡傪挷傋偨偲偙傠丄A兝偼塼徎憡偲儕僢僾儖僎儖憡偺PC枌偵偼寢崌偟傑偣傫偑丄儔儊儔僎儖憡偺PC枌偵懳偟偰偼寢崌偡傞偙偲偑暘偐傝傑偟偨乮恾係乯丅 儔儊儔僎儖憡偱側偗傟偽A兝偑寢崌偟側偄棟桼偼晄柧偱偡偑丄枌撪偱帀幙暘巕偑枾偵僷僢僉儞僌偝傟偰偄傞丄傑偨偼儕僢僾儖僎儖憡偲斾傋偰枌昞柺偑妸傜偐偱偁傞丄偲偄偆儔儊儔僎儖憡偺摿挜偑娭學偡傞偲梊憐偝傟傑偡丅 嵶朎枌偼僐儗僗僥儘乕儖傪娷傓偨傔丄PC偺傒偺儌僨儖帀幙枌偑掅壏偱偲傞傛偆側僎儖憡偺忬懺偵偼側傝傑偣傫丅偟偐偟丄拋彉塼懱憡偺忬懺偵偁傞偲峫偊傜傟傞帀幙儔僼僩偵偍偄偰偼丄帀幙暘巕偼斾妑揑崅偄棳摦惈傪曐偪偮偮傕儔儊儔僎儖憡偲摨條偵枾偵僷僢僉儞僌偝傟偰偄傞偲峫偊傜傟偰偄傑偡丅 枌傪峔惉偡傞帀幙暘巕偺僷僢僉儞僌偑儔僼僩撪奜偱堎側傞偙偲偑丄嵶朎枌偵寢崌偟偨A兝偑儔僼僩傊廤愊偡傞偙偲偺堦場偱偁傞壜擻惈偑偁傝傑偡丅
 |
| 恾係丏A兝偲儂僗僼傽僠僕儖僐儕儞枌偺憡屳嶌梡 |
A兝偲帀幙擇廳枌乮儕億僜乕儉乯偺崿崌梟塼偺墌曃岝擇怓惈僗儁僋僩儖傪應掕偡傞偲丄枌偵寢崌偟偨A兝偺擇師峔憿偑暘偐傝傑偡丅 儔儊儔僎儖憡偺拞偱傕斾妑揑崅壏偺忦審偱偼丄枌偵寢崌偟偨A兝偼儌僲儅乕偱兛僿儕僢僋僗傪宍惉偟傑偡丅 偟偐偟丄壏搙掅壓偵傛傝A兝偼枌寢崌忬懺偱暘巕娫夛崌偟丄兝僔乕僩傪宍惉偡傞偙偲偑暘偐傝傑偟偨乮恾係乯丅 慜揮堏壏搙埲壓偱儔儊儔僎儖憡偵側偭偨PC枌偼丄偝傜偵椻媝偝傟傞偲丄傛傝枾側僷僢僉儞僌傪帩偮枌偵抜奒揑偵曄壔偟丄嵟廔揑偵偼寢徎偵嬤偄僷僢僉儞僌傪帩偮僒僽僎儖憡偲屇偽傟傞忬懺偵揮堏偡傞偙偲偑抦傜傟偰偄傑偡丅 A兝偑兛僿儕僢僋僗傪宍惉偡傞偐丄傑偨偼暘巕娫夛崌偟偰兝僔乕僩傪宍惉偡傞偐偼丄傾儖僣僴僀儅乕昦偺敪徢傪寛掕晅偗傞偐傕偟傟側偄廳梫側億僀儞僩偱偡丅 帀幙枌偺僷僢僉儞僌忬懺偺嬐偐側堘偄偑丄偙偺暘婒揰偵偍偄偰曽岦傪寛傔傞傂偲偮偺梫慺偲側偭偰偄傞偺偐傕偟傟傑偣傫丅 帀幙枌偵傛傞儁僾僠僪偺擇師峔憿惂屼儊僇僯僘儉傪夝柧偡傞偨傔丄摉尋媶幒偱偼丄偝傜偵尋媶傪恑傔偰偄傑偡丅
傾儈儘僀僪兝(A兝1-40)儁僾僠僪偺傾儈儘僀僪慄堐壔偑傾儖僣僴僀儅乕昦偺尨場偲側傞偺偱偁傟偽丄慄堐壔慾奞嵻偼傾儖僣僴僀儅乕昦帯椕栻偲側傞壜擻惈偑偁傝傑偡丅 丂傾儈儘僀僪慄堐宍惉偼暘巕娫兝僔乕僩宍惉斀墳丄偡側傢偪儁僾僠僪庡嵔娫偱偺悈慺寢崌宍惉斀墳偱偡丅 偟偨偑偭偰丄宍惉慾奞嵻偵媮傔傜傟傞忦審偼(i) A兝1-40慄堐偵懳偡傞恊榓惈偑崅偄偙偲丄偍傛傃(ii)暘巕娫悈慺寢崌宍惉傪岠棪揑偵慾奞偡傞偙偲丄偱偡丅偙傟傜偺忦審傪枮偨偡暔幙偲偟偰丄変乆偼A兝1-40偺晹暘攝楍儁僾僠僪A兝30-40[攝楍30AIIGLMVGGVV40]傪慖傃丄庡嵔CONH婎傪悢巆婎偱儊僠儖壔偡傞乮CON(Me)婎偵抲偒姺偊傞乯偙偲偱慾奞嵻偲偟偰偺擻椡傪夝愅偟傑偟偨丅 慄堐宍惉傪旕忢偵岠棪傛偔慾奞偡傞暔幙傪愝寁偡傞偨傔偺巜恓傪摼偰丄岠壥揑側怢挿慾奞嵻傪嶌惢偡傞偙偲偑杮尋媶偺栚揑偱偡丅
N-儊僠儖壔巆婎偺堎側傞俉庬椶偺N-儊僠儖壔儁僾僠僪乮恾5乯100兪M傪A兝1-40 (50兪M)偵懳偟壛偊偨寢壥丄G37偍傛傃G38傪儊僠儖壔偟偨応崌偵慄堐宍惉検偑栺20%偵傑偱掅壓偡傞偙偲丄L34V36G38偺俁巆婎傪儊僠儖壔偟偨儁僾僠僪偱偼慄堐宍惉検偑77%偵偟偐側傜側偄偙偲丄偑柧傜偐偵側傝傑偟偨丅
N-儊僠儖壔巆婎傪摫擖偡傞偙偲偵傛傝悈慺寢崌宍惉傪慾奞偡傞偙偲傪堄恾偟傑偟偨偑丄慄堐壔慾奞擻偲N-儊僠儖壔巆婎偺悢偲偺娫偵偼憡娭偑尒傜傟偢丄N-儊僠儖壔傪峴偆巆婎偼侾巆婎偱廫暘偱偁傞丄偲偄偆寢壥傪摼傑偟偨丅
墌曃岝擇怓惈應掕偺寢壥偐傜丄N-儊僠儖壔儁僾僠僪偺僗儁僋僩儖偵尰傟傞220 nm偺晧偺僺乕僋偑尠挊偱偁傞応崌偵偼慄堐壔慾奞擻偑彫偝偔丄尠挊偱側偄応崌偵慾奞擻偑戝偒偄丄偲偄偆偙偲傪尒偄偩偟傑偟偨丅
偙偺僺乕僋偺嫮搙偼丄悈梟塼拞偱偺N-儊僠儖壔儁僾僠僪偺僐儞僼僅儊乕僔儑儞偲娭楢偡傞偲峫偊傜傟傞偙偲偐傜丄N-儊僠儖壔儁僾僠僪偺悈梟塼拞偱偺峔憿偑慄堐壔慾奞擻偵娭楢偡傞壜擻惈偑帵嵈偝傟傑偡丅
慄堐壔慾奞擻偺戝彫傪寛傔傞梫場偵娭偟偰丄尰嵼偝傜偵専摙傪恑傔偰偄傑偡丅
 |
| 恾俆丏梡偄偨N-儊僠儖壔A兝30-40偺攝楍 愒暥帤偼N-儊僠儖壔巆婎傪帵偡 |